
Tune in to the latest #eLifePodcast to hear authors discuss the stories behind their recent discoveries https://t.co/awwqu5L3lE
eLife - the journal (@eLife)
About Profile
[Do not use this base pattern directly: see the following variants of this pattern for usage]
About Profile Image

Randy Scheckman
Editor-in-Chief
Lorem ipsum dolor sit amet, consectetur adipiscing elit. Mauris tempor euismod mi. Morbi facilisis eros in mi consequat cursus. Nulla malesuada pretium mauris, vitae gravida diam laoreet ac. Curabitur ornare libero sed porta molestie. Ut ut sem in libero tincidunt lobortis. Nullam eros nisi, malesuada vitae leo sit amet, bibendum congue justo. Morbi iaculis lectus erat, vel dapibus metus posuere sit amet. Phasellus eget scelerisque massa. Mauris hendrerit ex in nisl ultricies elementum. Donec tellus risus, rhoncus molestie rutrum et, cursus eget mi. Praesent et elit tellus. Curabitur id sodales turpis. Fusce pulvinar aliquam commodo. Cras vitae dolor vitae metus dignissim auctor.Cras nulla purus, rhoncus et mollis in, imperdiet sed metus. In non porttitor nibh. In sit amet ipsum eget nisl laoreet tincidunt. Donec iaculis nibh quis lacus viverra sagittis. Phasellus imperdiet congue imperdiet.
Interdum et malesuada fames ac ante ipsum primis in faucibus. Nunc nisi tortor, rhoncus.
- Expertise
- Cell Biology
- Competing interests statement
- Donec tellus
About Profile No Image
Randy Scheckman
Editor-in-Chief
Lorem ipsum dolor sit amet, consectetur adipiscing elit. Mauris tempor euismod mi. Morbi facilisis eros in mi consequat cursus. Nulla malesuada pretium mauris, vitae gravida diam laoreet ac. Curabitur ornare libero sed porta molestie. Ut ut sem in libero tincidunt lobortis. Nullam eros nisi, malesuada vitae leo sit amet, bibendum congue justo. Morbi iaculis lectus erat, vel dapibus metus posuere sit amet. Phasellus eget scelerisque massa. Mauris hendrerit ex in nisl ultricies elementum. Donec tellus risus, rhoncus molestie rutrum et, cursus eget mi. Praesent et elit tellus. Curabitur id sodales turpis. Fusce pulvinar aliquam commodo. Cras vitae dolor vitae metus dignissim auctor.Cras nulla purus, rhoncus et mollis in, imperdiet sed metus. In non porttitor nibh. In sit amet ipsum eget nisl laoreet tincidunt. Donec iaculis nibh quis lacus viverra sagittis. Phasellus imperdiet congue imperdiet.
Interdum et malesuada fames ac ante ipsum primis in faucibus. Nunc nisi tortor, rhoncus.
- Expertise
- Cell Biology
- Competing interests statement
- Donec tellus
About Profile Placeholder Image

Randy Scheckman
Editor-in-Chief
Lorem ipsum dolor sit amet, consectetur adipiscing elit. Mauris tempor euismod mi. Morbi facilisis eros in mi consequat cursus. Nulla malesuada pretium mauris, vitae gravida diam laoreet ac. Curabitur ornare libero sed porta molestie. Ut ut sem in libero tincidunt lobortis. Nullam eros nisi, malesuada vitae leo sit amet, bibendum congue justo. Morbi iaculis lectus erat, vel dapibus metus posuere sit amet. Phasellus eget scelerisque massa. Mauris hendrerit ex in nisl ultricies elementum. Donec tellus risus, rhoncus molestie rutrum et, cursus eget mi. Praesent et elit tellus. Curabitur id sodales turpis. Fusce pulvinar aliquam commodo. Cras vitae dolor vitae metus dignissim auctor.Cras nulla purus, rhoncus et mollis in, imperdiet sed metus. In non porttitor nibh. In sit amet ipsum eget nisl laoreet tincidunt. Donec iaculis nibh quis lacus viverra sagittis. Phasellus imperdiet congue imperdiet.
Interdum et malesuada fames ac ante ipsum primis in faucibus. Nunc nisi tortor, rhoncus.
- Expertise
- Cell Biology
- Competing interests statement
- Donec tellus
Additional Asset
[Do not use this base pattern directly: see the following variants of this pattern for usage]
Additional Asset Doi
Table 1 - source data 1.
Is the species name mentioned in the title, impact statement, abstract and digest of eLife papers (July–Sept 2014)?
some more details here
Additional Asset Non Doi
Table 1 - source data 1.
Is the species name mentioned in the title, impact statement, abstract and digest of eLife papers (July–Sept 2014)?
some more details here
Article Download Links List
A three part list of links to download the article, or parts of the article, in various formats.
Downloads (links to download the article or parts of the article as PDF, and in DAR format)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa
eLife 4:e09560.
https://doi.org/10.7554/eLife.09560 Article Section Kitchen Sink
The Kitchen Sink
Some linkHeading 1
Heading 2
Heading 3
Heading 4
Heading 5
Heading 6
This is not wrapped in a paragraph.
This is in a paragraph. Lorem ipsum dolor sit amet, test link adipiscing elit. This is strong. Nullam dignissim convallis est. This is underlined. Quisque aliquam. This is emphasized. Donec faucibus. This is monospace. Nunc iaculis suscipit dui. 53 = 125. Water is H2O. Nam sit amet sem. This is in small caps. Aliquam libero nisi, imperdiet at, tincidunt nec, gravida vehicula, nisl. The New York Times (That’s a citation). Maecenas ornare tortor. Donec sed tellus eget sapien fringilla nonummy. Mauris a ante. Suspendisse quam sem, consequat at, commodo vitae, feugiat in, nunc. Morbi imperdiet augue quis tellus.
HTML and CSS are our tools. Mauris a ante. Suspendisse quam sem, consequat at, commodo vitae, TAATAAGGAAGAACTGCTTATTCTTAATTATTTCTACCTACTAAACTAACTAATTATCAACAAATATCATCTATTTAATAGTATATCATCACATGCGGTGTAAGAGGATGACATAAAGATTGAGAAACAGTCATCCAGTCTAATGGAAGCTCAAATGCAAGGGCTGATAATGTAATAGGATAATGAATGACAACGTATAAAAGGAAAGAAGATAAAGCAATATTATTTTGTAGAATTATCGATTCCCTTTTGTGGATCCCTATATCCTCGAGGAGAA feugiat in, nunc. Morbi imperdiet augue quis tellus. Praesent mattis, massa quis luctus fermentum, turpis mi volutpat justo, eu volutpat enim diam eget metus. To copy a file type COPY filename. Dinner’s at 5:00. Let’s make that 7. This text has been struck.
List Types
Definition List
- Definition List Title
- This is a definition list division.
- Definition
- An exact statement or description of the nature, scope, or meaning of something: our definition of what constitutes poetry.
Ordered List
- List Item 1
- List Item 2
- Nested list item A
- Nested list item B
- List Item 3
Unordered List
- List Item 1
- List Item 2
- Nested list item A
- Nested list item B
- List Item 3
Table
| Table Header 1 | Table Header 2 | Table Header 3 | Table Header 4 |
|---|---|---|---|
| Division 1 | Division 2 | Division 3 | Division 4 |
| Division 1 | Division 2 | Division 3 | Division 4 |
| Division 1 | Division 2 | Division 3 with link | Division 4 with link |
| This is blue. | This is green. | This is orange. | This is yellow. |
| This is purple. | This is red. | This is pink. | This is grey. |
Preformatted Text
Typographically, preformatted text is not the same thing as code. Sometimes, a faithful execution of the text requires preformatted text that may not have anything to do with code. Most browsers use Courier and that’s a good default — with one slight adjustment, Courier 10 Pitch over regular Courier for Linux users. For example:
“Beware the Jabberwock, my son! The jaws that bite, the claws that catch! Beware the Jubjub bird, and shun The frumious Bandersnatch!”
Code
Code can be presented inline, like <?php bloginfo('stylesheet_url'); ?>, or within a <pre> block. Because we have more specific typographic needs for code, we’ll specify Consolas and Monaco ahead of the browser-defined monospace font.
#container {
float: left;
margin: 0 -240px 0 0;
width: 100%;
}
Blockquotes
Let’s keep it simple. Italics are good to help set it off from the body text. Be sure to style the citation.
Good afternoon, gentlemen. I am a HAL 9000 computer. I became operational at the H.A.L. plant in Urbana, Illinois on the 12th of January 1992. My instructor was Mr. Langley, and he taught me to sing a song. If you’d like to hear it I can sing it for you.
— Citing HAL 9000
And here’s a bit of trailing text.
Article Section
[Do not use this base pattern directly: see the following variants of this pattern for usage]
Article Section Assessment
Assessment
This valuable paper informs on the role of type I PRMTs in programming muscle stem cell identification. The evidence presented is mostly solid, with some weaknesses in the evidence regarding the proposed mechanism. The paper will be of particular interest to those who study skeletal muscle satellite cell biology.
https://doi.org/10.7554/eLife.62927.sa1Significance of the findings:
Valuable: Contains findings that have theoretical or practical implications for a subfield.
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Solid: Methods, data and analyses broadly support the claims with only minor weaknesses.
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Article Section Default
Introduction
TNF Receptor Associated Factor 2 (TRAF2) is an adaptor protein that transduces signals following ligation of certain cytokine receptors including those binding TNF. It was first identified together with TRAF1 as a component of TNF receptor-2 and then TNF receptor-1 (TNFR1) signalling complexes (Rothe et al., 1994; Shu et al., 1996). TRAF2, like most other TRAFs, contains a RING domain, several zinc fingers, a TRAF-N, and a conserved TRAF-C domain which is responsible for oligomerisation and receptor binding through its MATH region (Takeuchi et al., 1996; Uren and Vaux, 1996).
RING domains are nearly always associated with ubiquitin E3 ligase activity (Shi and Kehrl, 2003) and TRAF2 can promote ubiquitylation of RIPK1 in TNFR1 signalling complexes (TNFR1-SC) (Wertz et al., 2004). However TRAF2 recruits E3 ligases such as cIAPs to TNFR1-SC and these have also been shown to be able to ubiquitylate RIPK1 and regulate TNF signalling (Dynek et al., 2010; Mahoney et al., 2008; Varfolomeev et al., 2008; Vince et al., 2009). This makes it difficult to unambiguously determine the role of the E3 ligase activity of TRAF2.
Activation of JNK and NF-κB by TNF is reduced in cells from Traf2-/- mice while only JNK signalling was affected in lymphocytes from transgenic mice that express a dominant negative (DN) form of TRAF2 that lacks the RING domain (Lee et al., 1997; Yeh et al., 1997). Traf2-/-Traf5-/- mouse embryonic fibroblasts (MEFs) have a pronounced defect in activation of NF-κB by TNF, suggesting that absence of TRAF2 can be compensated by TRAF5 (Tada et al., 2001). Although activation of NF-κB was restored in Traf2-/-Traf5-/- cells by re-expression of wild type TRAF2, it was not restored when the cells were reconstituted with TRAF2 point mutants that could not bind cIAPs (Vince et al., 2009; Zhang et al., 2010). These data, together with a wealth of different lines of evidence showing that cIAPs are critical E3 ligases required for TNF-induced canonical NF-κB (Blackwell et al., 2013; Haas et al., 2009; Silke, 2011), support the idea that the main function of TRAF2 in TNF-induced NF-κB is to recruit cIAPs to the TNFR1-SC. However, it remains possible that the RING of TRAF2 plays another function, such as in activating JNK and protecting cells from TNF-induced cell death (Vince et al., 2009; Zhang et al., 2010). Furthermore it has been shown that TRAF2 can K48-ubiquitylate caspase-8 to set the threshold for TRAIL or Fas induced cell death (Gonzalvez et al., 2012). Moreover, TRAF2 inhibits non-canonical NF-κB signalling (Grech et al., 2004; Zarnegar et al., 2008) and this function requires the RING domain of TRAF2 to induce proteosomal degradation of NIK (Vince et al., 2009). However, structural and in vitro analyses indicate that, unlike TRAF6, the RING domain of TRAF2 is unable to bind E2 conjugating enzymes (Yin et al., 2009), and is therefore unlikely to have intrinsic E3 ligase activity.
Sphingosine-1-phosphate (S1P) is a pleiotropic sphingolipid mediator that regulates proliferation, differentiation, cell trafficking and vascular development (Pitson, 2011). S1P is generated by sphingosine kinase 1 and 2 (SPHK1 and SPHK2) (Kohama et al., 1998; Liu et al., 2000). Extracellular S1P mainly acts by binding to its five G protein-coupled receptors S1P1-5 (Hla and Dannenberg, 2012). However, some intracellular roles have been suggested for S1P, including the blocking of the histone deacetylases, HDAC1/2 (Hait et al., 2009) and the induction of apoptosis through interaction with BAK and BAX (Chipuk et al., 2012).
Recently, it was suggested that the RING domain of TRAF2 requires S1P as a co-factor for its E3 ligase activity (Alvarez et al., 2010). Alvarez and colleagues proposed that SPHK1 but not SPHK2 is activated by TNF and phosphorylates sphingosine to S1P which in turn binds to the RING domain of TRAF2 and serves as an essential co-factor that was missing in the experiments of Yin et al. Alvarez and colleagues, observed that in the absence of SPHK1, TNF-induced NF-κB activation was completely abolished.
Although we know a lot about TRAF2, there are still important gaps particularly with regard to cell type specificity and in vivo function of TRAF2. Moreover, despite the claims that SPHK1 and its product, S1P, are required for TRAF2 to function as a ubiquitin ligase, the responses of Traf2-/- and Sphk1-/- cells to TNF were not compared. Therefore, we undertook an analysis of TRAF2 and SPHK1 function in TNF signalling in a number of different tissues.
Surprisingly, we found that neither TRAF2 nor SPHK1 are required for TNF mediated canonical NF-κB and MAPK signalling in macrophages. However, MEFs, murine dermal fibroblasts (MDFs) and keratinocytes required TRAF2 but not SPHK1 for full strength TNF signalling. In these cell types, absence of TRAF2 caused a delay in TNF-induced activation of NF-κB and MAPK, and sensitivity to killing by TNF was increased. Absence of TRAF2 in keratinocytes in vivo resulted in psoriasis-like epidermal hyperplasia and skin inflammation. Unlike TNF-dependent genetic inflammatory skin conditions, such as IKK2 epidermal knock-out (Pasparakis et al., 2002) and the cpdm mutant (Gerlach et al., 2011), the onset of inflammation was only delayed, and not prevented by deletion of TNF. This early TNF-dependent inflammation is caused by excessive apoptotic but not necroptotic cell death and could be prevented by deletion of Casp8. We observed constitutive activation of NIK and non-canonical NF-κB in Traf2-/- keratinocytes which caused production of inflammatory cytokines and chemokines. We were able to reverse this inflammatory phenotype by simultaneously deleting both Tnf and Nfkb2 genes. Our results highlight the important role TRAF2 plays to protect keratinocytes from cell death and to down-regulate inflammatory responses and support the idea that intrinsic defects in keratinocytes can initiate psoriasis-like skin inflammation.
https://doi.org/10.7554/eLife.16370Article Section Editor Evaluation
Editor's evaluation
The manuscript by Rosello et al. represents a major advance in implementation of cutting-edge genome editing methodologies in the zebrafish. The study seeks to describe optimized tools for precise base editing in zebrafish and to demonstrate their effective application. Overall, this study demonstrates that cytosine base editing is an efficient and powerful method for introducing precise in vivo edits into the zebrafish genome, and will be of interest to scientists who use zebrafish and other genetic systems to model human development and disease.
https://doi.org/10.7554/eLife.62927.sa1Article Section Editor Title Text
Senior editor
- Dominique Soldati-Favre
- University of Geneva, Switzerland
Article Section Header Link
TNF Receptor Associated Factor 2 (TRAF2) is an adaptor protein that transduces signals following ligation of certain cytokine receptors
some text to carry a linkTNF Receptor Associated Factor 2 (TRAF2) is an adaptor protein that transduces signals following ligation of certain cytokine receptors including those binding TNF. It was first identified together with TRAF1 as a component of TNF receptor-2 and then TNF receptor-1 (TNFR1) signalling complexes (Rothe et al., 1994; Shu et al., 1996). TRAF2, like most other TRAFs, contains a RING domain, several zinc fingers, a TRAF-N, and a conserved TRAF-C domain which is responsible for oligomerisation and receptor binding through its MATH region (Takeuchi et al., 1996; Uren and Vaux, 1996).
RING domains are nearly always associated with ubiquitin E3 ligase activity (Shi and Kehrl, 2003) and TRAF2 can promote ubiquitylation of RIPK1 in TNFR1 signalling complexes (TNFR1-SC) (Wertz et al., 2004). However TRAF2 recruits E3 ligases such as cIAPs to TNFR1-SC and these have also been shown to be able to ubiquitylate RIPK1 and regulate TNF signalling (Dynek et al., 2010; Mahoney et al., 2008; Varfolomeev et al., 2008; Vince et al., 2009). This makes it difficult to unambiguously determine the role of the E3 ligase activity of TRAF2.
Activation of JNK and NF-κB by TNF is reduced in cells from Traf2-/- mice while only JNK signalling was affected in lymphocytes from transgenic mice that express a dominant negative (DN) form of TRAF2 that lacks the RING domain (Lee et al., 1997; Yeh et al., 1997). Traf2-/-Traf5-/- mouse embryonic fibroblasts (MEFs) have a pronounced defect in activation of NF-κB by TNF, suggesting that absence of TRAF2 can be compensated by TRAF5 (Tada et al., 2001). Although activation of NF-κB was restored in Traf2-/-Traf5-/- cells by re-expression of wild type TRAF2, it was not restored when the cells were reconstituted with TRAF2 point mutants that could not bind cIAPs (Vince et al., 2009; Zhang et al., 2010). These data, together with a wealth of different lines of evidence showing that cIAPs are critical E3 ligases required for TNF-induced canonical NF-κB (Blackwell et al., 2013; Haas et al., 2009; Silke, 2011), support the idea that the main function of TRAF2 in TNF-induced NF-κB is to recruit cIAPs to the TNFR1-SC. However, it remains possible that the RING of TRAF2 plays another function, such as in activating JNK and protecting cells from TNF-induced cell death (Vince et al., 2009; Zhang et al., 2010). Furthermore it has been shown that TRAF2 can K48-ubiquitylate caspase-8 to set the threshold for TRAIL or Fas induced cell death (Gonzalvez et al., 2012). Moreover, TRAF2 inhibits non-canonical NF-κB signalling (Grech et al., 2004; Zarnegar et al., 2008) and this function requires the RING domain of TRAF2 to induce proteosomal degradation of NIK (Vince et al., 2009). However, structural and in vitro analyses indicate that, unlike TRAF6, the RING domain of TRAF2 is unable to bind E2 conjugating enzymes (Yin et al., 2009), and is therefore unlikely to have intrinsic E3 ligase activity.
Sphingosine-1-phosphate (S1P) is a pleiotropic sphingolipid mediator that regulates proliferation, differentiation, cell trafficking and vascular development (Pitson, 2011). S1P is generated by sphingosine kinase 1 and 2 (SPHK1 and SPHK2) (Kohama et al., 1998; Liu et al., 2000). Extracellular S1P mainly acts by binding to its five G protein-coupled receptors S1P1-5 (Hla and Dannenberg, 2012). However, some intracellular roles have been suggested for S1P, including the blocking of the histone deacetylases, HDAC1/2 (Hait et al., 2009) and the induction of apoptosis through interaction with BAK and BAX (Chipuk et al., 2012).
Recently, it was suggested that the RING domain of TRAF2 requires S1P as a co-factor for its E3 ligase activity (Alvarez et al., 2010). Alvarez and colleagues proposed that SPHK1 but not SPHK2 is activated by TNF and phosphorylates sphingosine to S1P which in turn binds to the RING domain of TRAF2 and serves as an essential co-factor that was missing in the experiments of Yin et al. Alvarez and colleagues, observed that in the absence of SPHK1, TNF-induced NF-κB activation was completely abolished.
Although we know a lot about TRAF2, there are still important gaps particularly with regard to cell type specificity and in vivo function of TRAF2. Moreover, despite the claims that SPHK1 and its product, S1P, are required for TRAF2 to function as a ubiquitin ligase, the responses of Traf2-/- and Sphk1-/- cells to TNF were not compared. Therefore, we undertook an analysis of TRAF2 and SPHK1 function in TNF signalling in a number of different tissues.
Surprisingly, we found that neither TRAF2 nor SPHK1 are required for TNF mediated canonical NF-κB and MAPK signalling in macrophages. However, MEFs, murine dermal fibroblasts (MDFs) and keratinocytes required TRAF2 but not SPHK1 for full strength TNF signalling. In these cell types, absence of TRAF2 caused a delay in TNF-induced activation of NF-κB and MAPK, and sensitivity to killing by TNF was increased. Absence of TRAF2 in keratinocytes in vivo resulted in psoriasis-like epidermal hyperplasia and skin inflammation. Unlike TNF-dependent genetic inflammatory skin conditions, such as IKK2 epidermal knock-out (Pasparakis et al., 2002) and the cpdm mutant (Gerlach et al., 2011), the onset of inflammation was only delayed, and not prevented by deletion of TNF. This early TNF-dependent inflammation is caused by excessive apoptotic but not necroptotic cell death and could be prevented by deletion of Casp8. We observed constitutive activation of NIK and non-canonical NF-κB in Traf2-/- keratinocytes which caused production of inflammatory cytokines and chemokines. We were able to reverse this inflammatory phenotype by simultaneously deleting both Tnf and Nfkb2 genes. Our results highlight the important role TRAF2 plays to protect keratinocytes from cell death and to down-regulate inflammatory responses and support the idea that intrinsic defects in keratinocytes can initiate psoriasis-like skin inflammation.
Article Section Process Block
Peer review process
This article was published via eLife's original peer review and publishing model, which was phased out in 2023. eLife's editors accepted this article for publication after peer review and was subject to mandatory revision requests before it was published.
History
- Version of Record published
- Preprint posted
- Preprint posted
- Preprint posted
- Sent for peer review
Article Section Without Title
TNF Receptor Associated Factor 2 (TRAF2) is an adaptor protein that transduces signals following ligation of certain cytokine receptors including those binding TNF. It was first identified together with TRAF1 as a component of TNF receptor-2 and then TNF receptor-1 (TNFR1) signalling complexes (Rothe et al., 1994; Shu et al., 1996). TRAF2, like most other TRAFs, contains a RING domain, several zinc fingers, a TRAF-N, and a conserved TRAF-C domain which is responsible for oligomerisation and receptor binding through its MATH region (Takeuchi et al., 1996; Uren and Vaux, 1996).
RING domains are nearly always associated with ubiquitin E3 ligase activity (Shi and Kehrl, 2003) and TRAF2 can promote ubiquitylation of RIPK1 in TNFR1 signalling complexes (TNFR1-SC) (Wertz et al., 2004). However TRAF2 recruits E3 ligases such as cIAPs to TNFR1-SC and these have also been shown to be able to ubiquitylate RIPK1 and regulate TNF signalling (Dynek et al., 2010; Mahoney et al., 2008; Varfolomeev et al., 2008; Vince et al., 2009). This makes it difficult to unambiguously determine the role of the E3 ligase activity of TRAF2.
Activation of JNK and NF-κB by TNF is reduced in cells from Traf2-/- mice while only JNK signalling was affected in lymphocytes from transgenic mice that express a dominant negative (DN) form of TRAF2 that lacks the RING domain (Lee et al., 1997; Yeh et al., 1997). Traf2-/-Traf5-/- mouse embryonic fibroblasts (MEFs) have a pronounced defect in activation of NF-κB by TNF, suggesting that absence of TRAF2 can be compensated by TRAF5 (Tada et al., 2001). Although activation of NF-κB was restored in Traf2-/-Traf5-/- cells by re-expression of wild type TRAF2, it was not restored when the cells were reconstituted with TRAF2 point mutants that could not bind cIAPs (Vince et al., 2009; Zhang et al., 2010). These data, together with a wealth of different lines of evidence showing that cIAPs are critical E3 ligases required for TNF-induced canonical NF-κB (Blackwell et al., 2013; Haas et al., 2009; Silke, 2011), support the idea that the main function of TRAF2 in TNF-induced NF-κB is to recruit cIAPs to the TNFR1-SC. However, it remains possible that the RING of TRAF2 plays another function, such as in activating JNK and protecting cells from TNF-induced cell death (Vince et al., 2009; Zhang et al., 2010). Furthermore it has been shown that TRAF2 can K48-ubiquitylate caspase-8 to set the threshold for TRAIL or Fas induced cell death (Gonzalvez et al., 2012). Moreover, TRAF2 inhibits non-canonical NF-κB signalling (Grech et al., 2004; Zarnegar et al., 2008) and this function requires the RING domain of TRAF2 to induce proteosomal degradation of NIK (Vince et al., 2009). However, structural and in vitro analyses indicate that, unlike TRAF6, the RING domain of TRAF2 is unable to bind E2 conjugating enzymes (Yin et al., 2009), and is therefore unlikely to have intrinsic E3 ligase activity.
Sphingosine-1-phosphate (S1P) is a pleiotropic sphingolipid mediator that regulates proliferation, differentiation, cell trafficking and vascular development (Pitson, 2011). S1P is generated by sphingosine kinase 1 and 2 (SPHK1 and SPHK2) (Kohama et al., 1998; Liu et al., 2000). Extracellular S1P mainly acts by binding to its five G protein-coupled receptors S1P1-5 (Hla and Dannenberg, 2012). However, some intracellular roles have been suggested for S1P, including the blocking of the histone deacetylases, HDAC1/2 (Hait et al., 2009) and the induction of apoptosis through interaction with BAK and BAX (Chipuk et al., 2012).
Recently, it was suggested that the RING domain of TRAF2 requires S1P as a co-factor for its E3 ligase activity (Alvarez et al., 2010). Alvarez and colleagues proposed that SPHK1 but not SPHK2 is activated by TNF and phosphorylates sphingosine to S1P which in turn binds to the RING domain of TRAF2 and serves as an essential co-factor that was missing in the experiments of Yin et al. Alvarez and colleagues, observed that in the absence of SPHK1, TNF-induced NF-κB activation was completely abolished.
Although we know a lot about TRAF2, there are still important gaps particularly with regard to cell type specificity and in vivo function of TRAF2. Moreover, despite the claims that SPHK1 and its product, S1P, are required for TRAF2 to function as a ubiquitin ligase, the responses of Traf2-/- and Sphk1-/- cells to TNF were not compared. Therefore, we undertook an analysis of TRAF2 and SPHK1 function in TNF signalling in a number of different tissues.
Surprisingly, we found that neither TRAF2 nor SPHK1 are required for TNF mediated canonical NF-κB and MAPK signalling in macrophages. However, MEFs, murine dermal fibroblasts (MDFs) and keratinocytes required TRAF2 but not SPHK1 for full strength TNF signalling. In these cell types, absence of TRAF2 caused a delay in TNF-induced activation of NF-κB and MAPK, and sensitivity to killing by TNF was increased. Absence of TRAF2 in keratinocytes in vivo resulted in psoriasis-like epidermal hyperplasia and skin inflammation. Unlike TNF-dependent genetic inflammatory skin conditions, such as IKK2 epidermal knock-out (Pasparakis et al., 2002) and the cpdm mutant (Gerlach et al., 2011), the onset of inflammation was only delayed, and not prevented by deletion of TNF. This early TNF-dependent inflammation is caused by excessive apoptotic but not necroptotic cell death and could be prevented by deletion of Casp8. We observed constitutive activation of NIK and non-canonical NF-κB in Traf2-/- keratinocytes which caused production of inflammatory cytokines and chemokines. We were able to reverse this inflammatory phenotype by simultaneously deleting both Tnf and Nfkb2 genes. Our results highlight the important role TRAF2 plays to protect keratinocytes from cell death and to down-regulate inflammatory responses and support the idea that intrinsic defects in keratinocytes can initiate psoriasis-like skin inflammation.
Assessment
Significance of the findings:
Valuable: Contains findings that have theoretical or practical implications for a subfield.
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Solid: Methods, data and analyses broadly support the claims with only minor weaknesses.
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Authors Details
Author details
Authors
- Lee R Berger

- Bernhard Zipfel
- University of the Witwatersrand, South Africa
- University of Wisconsin-Madison, United States
- Texas A&M University, United States
- Duke University, United States
- University of Zurich, Switzerland
- University of Arkansas, United States
- University of Kent, United Kingdom
- Max Planck Institute for Evolutionary Anthropology, Germany
- Mercyhurst University, United States
- New York University, United States
- New York Consortium in Evolutionary Primatology, United States
- Dartmouth College, United States
- University of Colorado Denver, United States
- Loughborough University, United Kingdom
- Tulane University, United States
- Lehman College, United States
- American Museum of Natural History, United States
- University of Cape Town, South Africa
- Museo Nacional de Ciencias Naturales, Spain
- Modesto Junior College, United States
- Louisiana State University, United States
- Nazarbayev University, Kazakhstan
- University of Missouri, United States
- University of Kentucky College of Medicine, United States
- Simon Fraser University, Canada
- Université de Montréal, Canada
- Australian National University, Australia
- Biology Department, Universidad Autònoma de Madrid, Spain
- Midwestern University, United States
- Liverpool John Moores University, United Kingdom
- University of Pisa, Italy
- Chaffey College, United States
- University of Johannesburg, South Africa
- George Washington University, United States
- University of Colorado School of Medicine, United States
- Croatian Natural History Museum, Croatia
- University of Iowa, United States
- Lincoln Memorial University, United States
- Smithsonian Institution, United States
Authors Between Three And Ten Authors
- University of the Witwatersrand, South Africa
- University of Wisconsin-Madison, United States
- Texas A&M University, United States
- Duke University, United States
- University of Zurich, Switzerland
- University of Arkansas, United States
- University of Kent, United Kingdom
- Max Planck Institute for Evolutionary Anthropology, Germany
- Mercyhurst University, United States
- New York University, United States
- New York Consortium in Evolutionary Primatology, United States
- Dartmouth College, United States
- University of Colorado Denver, United States
- Loughborough University, United Kingdom
- Tulane University, United States
- Lehman College, United States
- American Museum of Natural History, United States
- University of Cape Town, South Africa
- Museo Nacional de Ciencias Naturales, Spain
- Modesto Junior College, United States
- Louisiana State University, United States
- Nazarbayev University, Kazakhstan
- University of Missouri, United States
- University of Kentucky College of Medicine, United States
- Simon Fraser University, Canada
- Université de Montréal, Canada
- Australian National University, Australia
- Biology Department, Universidad Autònoma de Madrid, Spain
- Midwestern University, United States
- Liverpool John Moores University, United Kingdom
- University of Pisa, Italy
- Chaffey College, United States
- University of Johannesburg, South Africa
- George Washington University, United States
- University of Colorado School of Medicine, United States
- Croatian Natural History Museum, Croatia
- University of Iowa, United States
- Lincoln Memorial University, United States
- Smithsonian Institution, United States
Authors Less Then Three Authors
- University of the Witwatersrand, South Africa
- University of Wisconsin-Madison, United States
- Texas A&M University, United States
- Duke University, United States
- University of Zurich, Switzerland
- University of Arkansas, United States
- University of Kent, United Kingdom
- Max Planck Institute for Evolutionary Anthropology, Germany
- Mercyhurst University, United States
- New York University, United States
- New York Consortium in Evolutionary Primatology, United States
- Dartmouth College, United States
- University of Colorado Denver, United States
- Loughborough University, United Kingdom
- Tulane University, United States
- Lehman College, United States
- American Museum of Natural History, United States
- University of Cape Town, South Africa
- Museo Nacional de Ciencias Naturales, Spain
- Modesto Junior College, United States
- Louisiana State University, United States
- Nazarbayev University, Kazakhstan
- University of Missouri, United States
- University of Kentucky College of Medicine, United States
- Simon Fraser University, Canada
- Université de Montréal, Canada
- Australian National University, Australia
- Biology Department, Universidad Autònoma de Madrid, Spain
- Midwestern University, United States
- Liverpool John Moores University, United Kingdom
- University of Pisa, Italy
- Chaffey College, United States
- University of Johannesburg, South Africa
- George Washington University, United States
- University of Colorado School of Medicine, United States
- Croatian Natural History Museum, Croatia
- University of Iowa, United States
- Lincoln Memorial University, United States
- Smithsonian Institution, United States
Call To Action

Hear more about our new Editor-in-Chief, and the latest developments at eLife
Call To Action Needs Js Cookie Expires
Hear more about our new Editor-in-Chief, and the latest developments at eLife
Captioned Asset
[Do not use this base pattern directly: see the following variants of this pattern for usage]
Captioned Asset Iframe Google Map
Many features are conserved between the mammalian nephron and planarian protonephridia.
(A) Image of a young adult C. elegans and schematic depicting the twelve pairs of sensory neurons in the anterior amphid individuals carrying a virus with k deleterious mutations at its endemic equilibrium. class carrying the fewest number of deleterious mutations is defined as mutation class k = 0. Inset: variation in the basic reproductive rate of infected individuals (gray histogram) and variation in the net reproductive rate R of infected individuals (brown histogram) resulting from variation in the number of deleterious mutations carried by circulating viruses.
Captioned Asset Iframe
Many features are conserved between the mammalian nephron and planarian protonephridia.
(A) Image of a young adult C. elegans and schematic depicting the twelve pairs of sensory neurons in the anterior amphid individuals carrying a virus with k deleterious mutations at its endemic equilibrium. class carrying the fewest number of deleterious mutations is defined as mutation class k = 0. Inset: variation in the basic reproductive rate of infected individuals (gray histogram) and variation in the net reproductive rate R of infected individuals (brown histogram) resulting from variation in the number of deleterious mutations carried by circulating viruses.
Captioned Asset Image
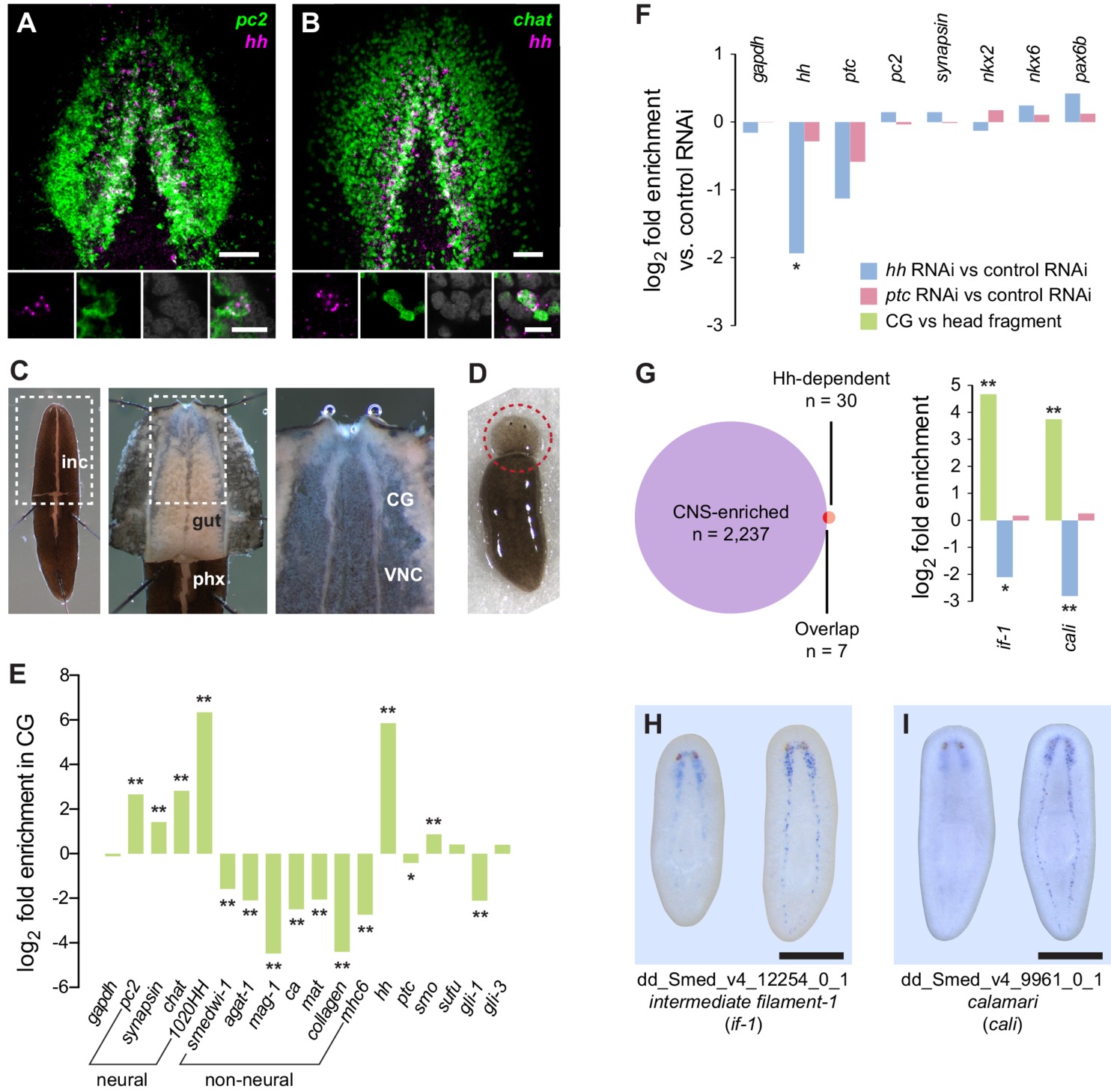
Many features are conserved between the mammalian nephron and planarian protonephridia.
(A) Image of a young adult C. elegans and schematic depicting the twelve pairs of sensory neurons in the anterior amphid individuals carrying a virus with k deleterious mutations at its endemic equilibrium. class carrying the fewest number of deleterious mutations is defined as mutation class k = 0. Inset: variation in the basic reproductive rate of infected individuals (gray histogram) and variation in the net reproductive rate R of infected individuals (brown histogram) resulting from variation in the number of deleterious mutations carried by circulating viruses.
Captioned Asset Inline Image
Many features are conserved between the mammalian nephron and planarian protonephridia.
(A) Image of a young adult C. elegans and schematic depicting the twelve pairs of sensory neurons in the anterior amphid individuals carrying a virus with k deleterious mutations at its endemic equilibrium. class carrying the fewest number of deleterious mutations is defined as mutation class k = 0. Inset: variation in the basic reproductive rate of infected individuals (gray histogram) and variation in the net reproductive rate R of infected individuals (brown histogram) resulting from variation in the number of deleterious mutations carried by circulating viruses.
Captioned Asset Picture

Many features are conserved between the mammalian nephron and planarian protonephridia.
(A) Image of a young adult C. elegans and schematic depicting the twelve pairs of sensory neurons in the anterior amphid individuals carrying a virus with k deleterious mutations at its endemic equilibrium. class carrying the fewest number of deleterious mutations is defined as mutation class k = 0. Inset: variation in the basic reproductive rate of infected individuals (gray histogram) and variation in the net reproductive rate R of infected individuals (brown histogram) resulting from variation in the number of deleterious mutations carried by circulating viruses.
Captioned Asset Table
Many features are conserved between the mammalian nephron and planarian protonephridia.
(A) Image of a young adult C. elegans and schematic depicting the twelve pairs of sensory neurons in the anterior amphid individuals carrying a virus with k deleterious mutations at its endemic equilibrium. class carrying the fewest number of deleterious mutations is defined as mutation class k = 0. Inset: variation in the basic reproductive rate of infected individuals (gray histogram) and variation in the net reproductive rate R of infected individuals (brown histogram) resulting from variation in the number of deleterious mutations carried by circulating viruses.
Captioned Asset Tables
Many features are conserved between the mammalian nephron and planarian protonephridia.
(A) Image of a young adult C. elegans and schematic depicting the twelve pairs of sensory neurons in the anterior amphid individuals carrying a virus with k deleterious mutations at its endemic equilibrium. class carrying the fewest number of deleterious mutations is defined as mutation class k = 0. Inset: variation in the basic reproductive rate of infected individuals (gray histogram) and variation in the net reproductive rate R of infected individuals (brown histogram) resulting from variation in the number of deleterious mutations carried by circulating viruses.
Captioned Asset Tweet
[Do not use this base pattern directly: see the following variants of this pattern for usage]
Captioned Asset Video
Many features are conserved between the mammalian nephron and planarian protonephridia.
(A) Image of a young adult C. elegans and schematic depicting the twelve pairs of sensory neurons in the anterior amphid individuals carrying a virus with k deleterious mutations at its endemic equilibrium. class carrying the fewest number of deleterious mutations is defined as mutation class k = 0. Inset: variation in the basic reproductive rate of infected individuals (gray histogram) and variation in the net reproductive rate R of infected individuals (brown histogram) resulting from variation in the number of deleterious mutations carried by circulating viruses.
Contextual Data
- Views 149
- Cited 5
- Annotations
Contextual Data With Citation
- Views 149
- Cited 5
- Annotations
Cite this article as: eLife 2016;5:e16370
doi:
10.7554/eLife.16370
Contextual Data With Download
- Views 149
- Downloads 20
- Cited 5
- Annotations

Highlight Item Blog Article

More eLife authors are linking submissions to their ORCID iDs
eLife sees positive results of requiring corresponding authors to register and link their profiles to their ORCID iDs
Highlight Item Event
eLife Continuum webinar
We're inviting all interested parties to participate in a webinar to learn about leveraging our new open-source publishing platform.
Highlight Item Press Pack
Scientists prove key aspect of evolutionary theory
While this event was first predicted almost twenty years ago, evidence for it has proved elusive.
Inline Profile

Lorem ipsum dolor sit amet, consectetur adipiscing elit. Mauris tempor euismod mi. Morbi facilisis eros in mi consequat cursus. Nulla malesuada pretium mauris, vitae gravida diam laoreet ac. Curabitur ornare libero sed porta molestie. Ut ut sem in libero tincidunt lobortis. Nullam eros nisi, malesuada vitae leo sit amet, bibendum congue justo. Morbi iaculis lectus erat, vel dapibus metus posuere sit amet. Phasellus eget scelerisque massa. Mauris hendrerit ex in nisl ultricies elementum. Donec tellus risus, rhoncus molestie rutrum et, cursus eget mi. Praesent et elit tellus. Curabitur id sodales turpis. Fusce pulvinar aliquam commodo. Cras vitae dolor vitae metus dignissim auctor.Cras nulla purus, rhoncus et mollis in, imperdiet sed metus. In non porttitor nibh. In sit amet ipsum eget nisl laoreet tincidunt. Donec iaculis nibh quis lacus viverra sagittis. Phasellus imperdiet congue imperdiet.
Interdum et malesuada fames ac ante ipsum primis in faucibus. Nunc nisi tortor, rhoncus.
Lorem ipsum dolor sit amet, consectetur adipiscing elit. Mauris tempor euismod mi. Morbi facilisis eros in mi consequat cursus. Nulla malesuada pretium mauris, vitae gravida diam laoreet ac. Curabitur ornare libero sed porta molestie. Ut ut sem in libero tincidunt lobortis. Nullam eros nisi, malesuada vitae leo sit amet, bibendum congue justo. Morbi iaculis lectus erat, vel dapibus metus posuere sit amet. Phasellus eget scelerisque massa. Mauris hendrerit ex in nisl ultricies elementum. Donec tellus risus, rhoncus molestie rutrum et, cursus eget mi. Praesent et elit tellus. Curabitur id sodales turpis. Fusce pulvinar aliquam commodo. Cras vitae dolor vitae metus dignissim auctor.Cras nulla purus, rhoncus et mollis in, imperdiet sed metus. In non porttitor nibh. In sit amet ipsum eget nisl laoreet tincidunt. Donec iaculis nibh quis lacus viverra sagittis. Phasellus imperdiet congue imperdiet.
Interdum et malesuada fames ac ante ipsum primis in faucibus. Nunc nisi tortor, rhoncus.





Media Chapter Listing Item
Drosophila duet: mating flies harmonise
Flies get to grips with acoustic duetting.
Mini Section
[heading]
[body]Personalised Cover Download
Download your personalised magazine cover
Thank you for publishing your research with us.
As a small keepsake for you and your co-authors we hope you’ll download this free poster highlighting your paper on an eLife magazine style cover.
Like your research it’s open access so you can print it, frame it, tweet it, use it as wallpaper in your lab...the possibilities are endless.
Profile Snippet
Name McNamerson
Title McTitleson
E: test.email@testemail.com
Pull Quote
Quote Incididunt ut labore et dolore magna aliqua. Ut enim ad minim veniam, quis nostrud exercitation ullamco laboris nisi ut reprehenderit in voluptate velit esse cillum dolore eu fugiat nulla pariatur.
Excepteur sint est laborum.
Pull Quote With Cite
Quote Incididunt ut labore et dolore magna aliqua. Ut enim ad minim veniam, quis nostrud exercitation ullamco laboris nisi ut reprehenderit in voluptate velit esse cillum dolore eu fugiat nulla pariatur. Excepteur sint est laborum.
May or may not be a link
Quote
Lorem ipsum dolor sit amet, consectetur adipiscing elit. Aenean interdum odio et sem fringilla, sit amet vulputate turpis sagittis. Cras non urna quam. Mauris vitae dui id ipsum varius semper. Curabitur sit amet viverra eros, ut maximus nisl. In hendrerit convallis turpis quis pulvinar. Donec porttitor faucibus ex, eget finibus velit vestibulum vitae. Donec nec nulla sollicitudin, volutpat enim interdum, sollicitudin erat.
Curabitur fermentum tellus ac rutrum efficitur. Phasellus libero magna, efficitur rutrum mauris a, mollis convallis lectus. Sed dapibus placerat eros ut vulputate. Vivamus quis sem accumsan, laoreet enim vel, dapibus ante. Nunc bibendum faucibus metus. Aliquam volutpat dapibus erat, quis ullamcorper nisl fermentum a. Praesent vulputate neque leo, vel rutrum dui auctor in. Duis eu magna et sem ullamcorper vehicula.
Quote With Cite
Lorem ipsum dolor sit amet, consectetur adipiscing elit. Aenean interdum odio et sem fringilla, sit amet vulputate turpis sagittis. Cras non urna quam. Mauris vitae dui id ipsum varius semper. Curabitur sit amet viverra eros, ut maximus nisl. In hendrerit convallis turpis quis pulvinar. Donec porttitor faucibus ex, eget finibus velit vestibulum vitae. Donec nec nulla sollicitudin, volutpat enim interdum, sollicitudin erat.
Curabitur fermentum tellus ac rutrum efficitur. Phasellus libero magna, efficitur rutrum mauris a, mollis convallis lectus. Sed dapibus placerat eros ut vulputate. Vivamus quis sem accumsan, laoreet enim vel, dapibus ante. Nunc bibendum faucibus metus. Aliquam volutpat dapibus erat, quis ullamcorper nisl fermentum a. Praesent vulputate neque leo, vel rutrum dui auctor in. Duis eu magna et sem ullamcorper vehicula.
May or may not be a link
Reference List
-
BookUnderstanding the rhythm of breathing: so near, yet so farIn: Julius D, Clapham DE, editors. Annual Review of Physiology 75:423–452.https://doi.org/10.1146/annurev-physiol-040510-130049
-
BookPhilosophie AnatomiqueParis: Méquignon-Marvis
-
Clinical TrialScripps Wired for Health Study (Registration: 000000)Scripps Translational Science Institute (sponsor) (2015)
-
ConferenceBoundary learning by optimization with topological constraintsBoundary learning by optimization with topological constraints, IEEE Conference on Computer Vision and Pattern Recognition (CVPR)https://doi.org/10.1038/365833a0
-
ConferenceBoundary learning by optimization with topological constraintsBoundary learning by optimization with topological constraints, IEEE Conference on Computer Vision and Pattern Recognition (CVPR)
-
ConferenceBoundary learning by optimization with topological constraintsBoundary learning by optimization with topological constraints, IEEE Conference on Computer Vision and Pattern Recognition (CVPR)
-
DataNKX2-5 mutations causative for congenital heart disease retain functionality and are directed to hundreds of targetsNCBI Gene Expression Omnibus. Series accession number GSE44902
-
PatentIRE-1alpha inhibitors (US20100941530)Mankind Corp, United States patent, United States
-
SoftwareThe Pymol Molecular Graphics SystemDeLano Scientific, Palo Alto, CA
Section Listing
Research categories
- Biochemistry
- Biophysics and structural biology
- Cancer biology
- Cell biology
- Computational and systems biology
- Developmental biology and stem cells
- Ecology
- Epidemiology and Global Health
- Genes and chromosomes
- Genomics and evolutionary biology
- Immunology
- Medicine
- Microbiology and infectious disease
- Neuroscience
- Plant biology
Section Listing Single
Magazine sections
Back to top
Sort Control
Sort by:
Statistic
- 320
- Downloads